Споровые растения: примеры, особенности, разновидности. Споровые растения
Общая характеристика растений. Споровые растения
Царство растения – Plantae, Vegetabilia
Общая характеристика царства
Представители царства – высокоспециализированные автотрофные организмы, питающиеся за счет процесса аэробного фотосинтеза. Их тело обычно расчленено на стебель, корень, лист, и они хорошо приспособлены к жизни в наземно-воздушной среде. Клетки растений имеют плотную клеточную стенку, в основе которой – целлюлоза. Основной запасной продукт – крахмал. Размножение вегетативное, бесполое (спорами) и половое (оогамия); мужские гаметы либо имеют ундулиподии (сперматозоиды), либо не имеют их (спермии). Характерно чередование полового (гаметофит) и бесполого поколения (спорофит) с преобладанием диплоидного бесполого поколения. Зигота у растений дает начало зародышу, который затем развивается в спорофит.
Царство растений включает не менее 300 тыс. видов (ныне существующих и вымерших), относящихся к 9 отделам – риниофиты (Rhyniophyta ) и зостерофиллофиты (Zosterophyllophyta ) (ныне вымершие), моховидные (Btyophyta), плауновидные (Lycopodiophyta ), псилотовидные (Psilotophyta ), хвощевидные (Equisetophyta ), папоротниковидные (Polypodiophyta ), голосеменные (Pinophyta) и покрытосеменные (Magnoliophyta ). Представители ныне существующих отделов, за исключением моховидных, характеризуются преобладанием в цикле развития бесполого поколения (спорофита), имеющего сосуды и (или) трахеиды. Благодаря последнему обстоятельству эти растения называют сосудистыми.
Растения делят на две группы: споровые и семенные . У споровых растений спорогенез и гаметогенез разобщены во времени и пространстве: спорофиты и гаметофиты являются отдельными физиологически самостоятельными организмами. Единицей размножения являются споры. У семенных растений гаметофиты сильно редуцированы и не являются физиологически самостоятельными организмами. Единицей размножения является семя.
Споровые растения являются первопоселенцами суши, которые дали начало в процессе эволюции семенным растениям.
Споровые растения
Включают следующие ныне существующие отделы: моховидные (Bryophyta ), плауновидные (Lycopodiophyta ), псилотовидные (Psilotophyta ), хвощевидные (Equisetophyta ), папоротниковидные (Polypodiophyta ).
Споровые растения появились в конце силурийского периода, более 400 млн. лет тому назад. Первые представители споровых были небольших размеров и имели простое строение, но уже у примитивных растений наблюдалась дифференциация на элементарные органы. Усовершенствованию органов соответствовало усложнение внутреннего строения и онтогенеза. В жизненном цикле происходит чередование полового и бесполого способов размножения и связанное с этим чередование поколений. Бесполое поколение представлено диплоидным спорофитом , половое – гаплоидным гаметофитом .
На спорофите образуютсяспорангии, внутри которых в результате мейотического деления образуются гаплоидные споры. Это мелкие, одноклеточные образования, лишенные жгутиков. Растения, у которых все споры одинаковые, называютсяравноспоровыми. У более высокоорганизованных групп споры двух типов:микроспоры (формируются в микроспорангиях), мегаспоры (формируются в мегаспорангиях). Это разноспоровые растения. При прорастании споры образуетсягаметофит.
Полный жизненный цикл (от зиготы до зиготы) состоит из гаметофита (период от споры до зиготы) испорофита (период от зиготы до образования спор).У плаунов, хвощей и папоротников эти фазы представляют собой как бы отдельные физиологически самостоятельные организмы.У мхов гаметофит является самостоятельной фазой жизненного цикла, а спорофит сведен до его своеобразного органа –спорогона (спорофит живет на гаметофите).
На гаметофите развиваются органы полового размножения:архегонии иантеридии . Вархегониях , похожих на колбу, образуются яйцеклетки, а в мешковидныхантеридиях - сперматозоиды. У равноспоровых растений гаметофиты обоеполые, у разноспоровых – однополые. Оплодотворение происходит лишь при наличии воды. При слиянии гамет образуется новая клетка – зигота с двойным набором хромосом (2n).
Отдел Моховидные – Bryophyta
Насчитывает до 27000 видов. Моховидные имеют тело либо в виде слоевища, либо расчленены на стебель и листья. Настоящих корней у них нет, их заменяют ризоиды. Проводящие ткани появляются лишь у высокоразвитых мхов. Частично обособлены ассимиляционная и механическая ткани.
В жизненном цикле доминирует гаметофит. Спорофит самостоятельно не существует, развивается и всегда находится на гаметофите, получая от него воду и питательные вещества. Спорофит представляет собой коробочку, где развивается спорангий, на ножке, связывающей ее с гаметофитом.
Размножаются мхи спорами, могут размножаться и вегетативным путем – отдельными участками тела или особыми выводковыми почками.
Отдел делится на три класса : Антоцеротовые (100 видов, шесть родов талломных растений), Печеночные и Листостебельные мхи.
Класс Печеночные мхи (Hepaticopsida )
Класс насчитывает около 8500 видов. Это, в основном, слоевищные мхи, хотя есть виды, имеющие стебель и листья. Широко распространена маршанция обыкновенная (Marchantia polymorpha )(рис 11. 1).
Рис. 11. 1. Цикл воспроизведения маршанции: 1 – таллом с мужскими подставками;2 – таллом с женскими подставками;3 – вертикальный разрез через мужскую подставку (в некоторых антеридиальных полостях находятся антеридии);4 – антеридий в антеридиальной полости (н – ножка антеридия);5 – двужгутиковый сперматозоид;6 – вертикальный разрез через женскую подставку (а – архегоний).
Гаметофит имеет темно-зеленоеслоевище (таллом), дихотомически разветвленное на широкие лопастные пластинки с дорсовентральной (спинно-брюшной) симметрией. Сверху и снизу слоевище покрыто эпидермой, внутри имеются ассимиляционная ткань и клетки, выполняющие проводящую и запасающую функции. К субстрату слоевище прикрепляетсяризоидами . На верхней стороне слоевища в особых «корзиночках» образуются выводковые почки, служащие для вегетативного размножения.
Талломы раздельнополые, органы полового размножения развиваются на особых вертикальных ветвях-подставках.
Мужские гаметофиты имеют восьмилопастные подставки, на верхней стороне которых находятся антеридии . На женских гаметофитах подставки со звездчатыми дисками, на нижней стороне лучей звездочки расположены (шейкой вниз)архегонии. При наличии воды сперматозоиды перемещаются, попадают в архегоний и сливаются с яйцеклеткой.
После оплодотворения из зиготы развивается спорогон. Он имеет вид шаровидной коробочки на короткой ножке. Внутри коробочки, в результате мейоза, из спорогенной ткани, образуются споры. В благоприятных условиях споры прорастают, из них развивается протонема в виде небольшой нити, из верхушечной клетки которой развивается таллом маршанции.
Класс Листостебельные мхи (Bryopsida, или Musci ).
Листостебельные мхи распространены по всему земному шару, особенно в условиях холодного климата на сырых местах, в сосновых и еловых лесах, в тундре. На торфяных и моховых болотах часто образуют плотный ковер. Тело расчленено на стебель и листья, но настоящих корней нет, есть многоклеточные ризоиды. Класс состоит из трех подклассов: Бриевые, или Зеленые мхи; Сфагновые, или Белые мхи; Андреевые, или Черные мхи.
Андреевые мхи (три рода, 90 видов) распространены в холодных областях, внешне сходны с зелеными, по строению листьев и коробочки – со сфагновыми.
Подкласс Бриевые, или Зеленые мхи (Bryidae ). Насчитывает около 700 родов, объединяющих 14000 видов, широко распространенных повсюду, особенно в тундровой и лесной зонах Северного полушария.
Широко распространен кукушкин лен (Polytrichium commune ), образующий густые дерновины на сырых почвах в лесах, на болотах и лугах. Стебли до40 см высоты, неветвистые, с густыми жесткими и острыми листьями. От нижней части стебля отходят ризоиды.
Цикл развития кукушкина льна (рис. 11. 2).

Рис. 11. 2.Кукушкин лен:А – цикл развития мха;Б – коробочка: 1 – с колпачком, 2 – без колпачка, 3 – в разрезе (а –крышечка, б – урночка, в – спорангий, г – апофиза, д – ножка);В – поперечный разрез листа с ассимиляторами;Г – поперечный разрез стебля (ф – флоэма, крв – крахмалоносное влагалище, кор – кора, э – эпидерма, лс – листовые следы).
Гаметофиты кукушкина льна раздельнополые. Рано весной на верхушке мужских особей развиваются антеридии, на верхушках женских – архегонии.
Весной во время дождя или после росы сперматозоиды выходят из антеридия и проникают в архегоний, где сливаются с яйцеклеткой. Из зиготы здесь же, на верхушке женского гаметофита, вырастает спорофит (спорогон), имеющий вид коробочки на длинной ножке. Коробочка покрыта волосистым колпачком (калиптра) (остатком архегония). В коробочке – спорангий, где после мейоза образуются споры. Спора – небольшая клетка с двумя оболочками. На верхушке коробочки, по ее краю, расположены зубцы (перистом), которые, в зависимости от влажности воздуха, загибаются внутрь коробочки или отгибаются наружу, чем способствуют рассеиванию спор. Споры разносятся ветром и в благоприятных условиях прорастают, образуя протонему. Через некоторое время на протонеме образуются почки, из которых формируются листостебельные побеги. Эти побеги вместе с протонемой – гаплоидное поколение – гаметофит. Коробочка на ножке – диплоидное поколение – спорофит.
Подкласс Сфагновые, или Белые мхи (Sphagnidae)
К сфагновым мхам относятся свыше 300 видов единственного рода сфагнум (Sphagnum )(рис. 11. 3).

Рис 11. 3.Сфагнум : 1 – внешний вид; 2 – верхушка ветви со спорогоном; 3 – спорогон (ш – остаток шейки архегония, кр – крышечка, сп – спорангий, кол – колонка, н – ножка спорогона, лн – ложная ножка); 4 – часть веточного листа (хлк – хлорофиллоносные клетки, вк – водоносные клетки, п – поры); 5 – поперечный разрез листа.
Ветвистые стебли сфагнума усеяны мелкими листьями. На верхушке главной оси боковые веточки образуют розетку почковидной формы. Особенностью сфагновых мхов является непрерывное нарастание стебля верхушкой и отмирание нижней части. Ризоиды отсутствуют, и поглощение воды с минеральными веществами происходит стеблями. Листья этих мхов состоят из двух типов клеток: 1) живых ассимилирующих, длинных и узких, хлорофиллоносных; 2) гиалиновых – мертвых, лишенных протопласта. Гиалиновые клетки легко заполняются водой и долго ее сохраняют. Благодаря такому строению, сфагновые мхи могут накапливать воды в 37 раз больше своей сухой массы. Разрастаясь плотными дернинами, сфагновые мхи способствуют заболачиванию почв. На болотах наслаивание отмерших частей мха приводит к образованию торфяников. Путем сухой перегонки из торфа получают воск, парафин, фенолы, аммиак; путем гидролиза – спирт. Торфяные плиты являются хорошим теплоизоляционным материалом. Сфагновые мхи обладают бактерицидными свойствами.
Отдел Плауновидные – Lycopodiophyta
Появление плауновидных связывают с силурийским периодом палеозойской эры. В настоящее время отдел представлен травянистыми растениями со стелющимися, дихотомически ветвящимися стеблями и корнями, а также спирально расположенными чешуйчатыми листьями. Листья возникли как выросты на стебле, и их называют микрофиллы . У плаунов есть флоэма, ксилема, перицикл.
Выделяют два современных класса: равноспоровые Плауновые и разноспоровые Полушниковые.
Класс Плауновые (Lycopodiopsida)
Из всего класса до настоящего времени сохранилось четыре рода.
Род плаун (Lycopodium). К этому роду относят многочисленные (около 200 видов) многолетние вечнозеленые травы, распространенные от арктических областей до тропиков. Так плаун булавовидный(L.clavatum) встречается в травяном покрове хвойных лесов на достаточно влажных, но бедных гумусом почвах. В сырых хвойных лесах широко распространен плаун годичный (L. annotinum )(рис. 11. 4).

Рис. 11. 4.Плаун булавовидный.
Родбаранец (Huperzia). Представитель рода- баранец обыкновенный (H. selago) распространен в тундровой, лесотундровой и северной лесной зонах и произрастает в южно-таежных еловых лесах и ольшаниках, а также в мшистых лесах и на альпийских лугах.
Роддифазиаструм (Diphasiastrum ). Представитель рода дифазиаструм сплюснутый(D. complanatum) растет на сухих песчаных почвах в сосновых лесах.
Цикл развития на примере плауна булавовидного (рис. 11. 5).

Рис. 11. 5. Цикл развития плауна булавовидного :1 – спорофит; 2 – спорофилл со спорангием; 3 – спора; 4 – гаметофит с антеридиями и архегониями; 5 – развивающийся на гаметофите из зародыша молодой спорофит.
Стелющиеся побеги плауна булавовидного достигают до 25 см в высоту и более 3 м в длину. Стебли покрыты спирально расположенными ланцетно-линейными мелкими листьями. В конце лета на боковых побегах образуется обычно два спороносных колоска. Каждый колосок состоит из оси и мелких тонких спорофиллов – видоизмененных листьев, в основании которых расположены спорангии почковидной формы.
В спорангиях после редукционного деления клеток спорогенной ткани образуются одинаковой величины, одетые толстой желтой оболочкой гаплоидныеспоры. Они прорастают после периода покоя через 3-8 лет в обоеполые заростки, представляющие собой половое поколение и живущиесапротрофно в почве, в виде клубенька. От нижней поверхности отходят ризоиды. Через них в заросток врастают гифы гриба, образуямикоризу . В симбиозе с грибом, который обеспечивает питание, живет заросток, лишенный хлорофилла и неспособный к фотосинтезу. Заростки многолетние, развиваются очень медленно, лишь через 6-15 лет на них образуются архегонии и антеридии. Оплодотворение совершается при наличии воды. После оплодотворения яйцеклетки двужгутиковым сперматозоидом образуется зигота, которая без периода покоя прорастает в зародыш, развивающийся во взрослое растение.
В официнальной медицине споры плауновприменялись в качестве детской присыпки и обсыпки для пилюль. Побеги баранца обыкновенного используют для лечения больных, страдающих хроническим алкоголизмом.
Класс Полушниковые (Isoetopsida)
Селагинелла (Selaginella ) среди современных родов насчитывает самое большое (около 700) число видов.
Это нежное многолетнее травянистое растение, требующее высокой влажности. Селагинеллы в отличие от плаунов характеризуются разноспоровостью. В спороносных колосках образуется два вида спор – четыремегаспоры в мегаспорангиях и многочисленныемикроспоры в микроспорангиях. Из микроспоры образуется мужской гаметофит, состоящий из одной ризоидальной клетки и антеридия со сперматозоидами. Мегаспора развивается в женский гаметофит, не покидающий ее оболочки и состоящий из мелкоклеточной ткани, в которую погружены архегонии. После оплодотворения из яйцеклетки развивается зародыш, и затем новый спорофит.
Отдел Хвощевидные – Equisetophyta
Появились хвощи в верхнем девоне, достигли наибольшего разнообразия в карбоне, когда древесный ярус заболоченных тропических лесов в значительной степени состоял из древовидных хвощевидных, вымерших к началу мезозоя. Современные хвощевидные появились на Земле с мелового периода.
До настоящего времени сохранился лишь один род – хвощ (Equisetum), представленный 30-35 видами, распространенными на всех континентах.
У всех видов хвоща стебли имеют членистое строение с выраженным чередованием узлов и междоузлий. Листья редуцированы до чешуй и располагаются мутовками в узлах. Здесь же образуются и боковые ветви. Ассимилирующую функцию выполняют зеленые стебли, поверхность которых увеличивается ребристостью, стенки клеток эпидермы пропитаны кремнеземом. Подземная часть представлена сильно развитым корневищем, в узлах которого формируются придаточные корни. У хвощаполевого (Equisetum arvense) боковые ветви корневища служат местом отложения запасных веществ, а также органами вегетативного размножения(рис. 11. 6).
<< < 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 > >>
Яндекс.Директ

Рис. 11. 6.Хвощ полевой : а, б – вегетативный и спороносный побеги спорофита; в – спорангиофор со спорангиями; г, д – споры; е – мужской гаметофит с антеридиями; ж – сперматозоид; з – обоеполый гаметофит; и - архегонии.
Весной на обычных или специальных спороносных стеблях образуются колоски, состоящие из оси, которая несет особые структуры, имеющие вид шестигранных щитков (спорангиофоры ). Последние несут 6-8 спорангиев. Внутри спорангиев образуются споры, одетые толстой оболочкой, снабженной гигроскопичными лентовидными выростами –элатерами. Благодаряэлатерам споры сцепляются вместе в комки, хлопья. Групповое распространение спор способствует тому, что при их прорастании разнополые заростки оказываются рядом, и это облегчает оплодотворение.
Заростки имеют вид небольшой длинно-лопастной зеленой пластинки с ризоидами на нижней поверхности. Мужские заростки меньше женских и несут по краям лопастей антеридии с многожгутиковыми сперматозоидами. На женских заростках в средней части развиваются архегонии. Оплодотворение происходит при наличии воды. Из зиготы развивается зародыш нового растения – спорофита.
В настоящее время большой роли в образовании растительного покрова хвощи не имеют. В лесах, на избыточно увлажненной почве, широко распространен хвощ лесной (E. sylvaticum) с сильно ветвящимися, поникающими боковыми веточками. На лугах, паровых полях, в посевах встречается трудно искореняемый сорнякхвощ полевой (E. arvense). У этого хвоща рано весной появляются неветвящиеся побеги, несущие спороносные колоски. Позднее от корневища развиваются зеленые вегетативные побеги. В лесной зоне на песчаных почвах и в оврагах широко распространенхвощ зимующий (E. hyemale ).
Вегетативные побеги хвоща полевого (E. arvense) в официнальной медицине применяют: в качестве мочегонного средства при отеках на почве сердечной недостаточности; при заболеваниях мочевого пузыря и мочевыводящих путей; как кровоостанавливающее средство при маточных кровотечениях; при некоторых формах туберкулеза.
Отдел Папоротниковидные – Polypodiophyta
Возникли папоротники в девоне, когда древовидные папоротники вместе с ныне ископаемыми плаунами и хвощами господствовали в растительном покрове земли. Большинство их вымерло, оставшиеся дали начало мезозойским формам, которые были представлены очень широко. Папоротники намного превосходят по числу современных видов все остальные отделы высших споровых (около 25000).
У большинства живущих сейчас (исключая тропические) папоротников отсутствует наземный прямостоячий стебель, но есть подземный в виде корневища. От корневища отходят придаточные корни и крупные листья (вайи ), имеющие стеблевое происхождение и длительно нарастающие верхушкой. Молодые листья обычно свернуты «улиткой». Среди существующих в настоящее время папоротников имеются какравноспоровые, так иразноспоровые.
В лесах Австралии, Южной Америки, Азии произрастают древовидные представители с колонновидными, неветвящимися, до 20 метров высоты стволами. В средней полосе нашей страны папоротники – это многолетние корневищные травы. Многие папоротники, как и мхи, являются индикаторами почв и типов леса. В светлых лесах, на песчаных или сухих подзолистых почвах распространен орляк обыкновенный (Pteridium aquilinum); на влажных богатых почвахкочедыжники (Athyrium) и крупныелесныещитовники (Dryopteris) (рис. 11. 7).

Рис. 11. 7.Щитовник мужской:А – спорофит: а – общий вид; б – сорусы на нижней стороне вайи; в – разрез соруса (1 – индузий, 2 – планцента, 3 – спорангий); г – спорангий (4 – кольцо);Б – гаметофит: 5 – сперматозоиды; 6 – заросток с нижней стороны (т – таллом, р – ризоиды, арх - архегонии, ан – антеридии); 7 – выход сперматозоидов из антеридия; 8 – архегоний с яйцеклеткой.
Цикл развития равноспоровых папоротников
В середине лета на нижней стороне зеленых листьев (у некоторых на специальных спороносных листьях) в виде коричневых бородавочек появляются группы спорангиев (сорусы) . Сорусы у многих папоротников прикрыты сверху своеобразным покрывалом -индузием. Спорангии образуются на особом выросте листа (плаценте) и имеют чечевицеобразную форму, длинные ножки и многоклеточные стенки. В спорангиях хорошо выражено механическое кольцо, которое имеет вид узкой несмыкающейся полосы, опоясывающей спорангий. Когда кольцо подсыхает, стенки спорангия разрываются, и споры высыпаются.
Споры, образовавшиеся в спорангиях, одноклеточны и имеют толстую оболочку. При созревании они разносятся током воздуха и в благоприятных условиях прорастают, образуя сердцевидную зеленую многоклеточную пластинку (заросток), прикрепляющуюся к почве ризоидами. Заросток представляет собой половое поколение папоротников (гаметофит). На нижней стороне заростка образуются антеридии (со сперматозоидами) и архегонии (с яйцеклетками). При наличии воды сперматозоиды проникают в архегонии и оплодотворяют яйцеклетки. Из зиготы развивается зародыш, имеющий все основные органы (корень, стебель, лист и особый орган – ножку, прикрепляющую его к заростку). Постепенно зародыш начинает существовать самостоятельно, а заросток отмирает.
У разноспоровых папоротников гаметофиты редуцированы до микроскопических размеров (особенно мужские).
Из корневищпапоротника мужского (Dryopteris filix-mas), получают густой экстракт, который является эффективным противоглистным средством (ленточные черви).
Красные книги - это списки редких и находящихся под угрозой исчезновения организмов; аннотированный перечень видов и подвидов с указанием прошлого и современного распространения, особенностей воспроизводства, уже принятых и необходимых мер по их охране. Различают международные, национальные и локальные Красные книги, а также отдельно Красные книги растений, животных и других систематических групп. Сбор информации для Красных книг был начат Международным союзом охраны природы и природных ресурсов (МСОП) в 1949 г.
Кроме Красной книги МСОП (1997 г.), перечень охраняемых видов растений и грибов в международном масштабе содержатся в Приложениях 1 и 2 Конвенции о международной торговле видами дикой фауны и флоры, находящимися под угрозой исчезновения (СИТЕС, 1973 г.).
Первое издание Красной книги РСФСР вышло в 80-х годах (том «Животные», 1983 г.; том «Растения», 1988 г.). В 2001 г. опубликовано новое издание Красной книги Российской Федерации, том «Животные».
Кроме Красных книг существую так называемые Зеленые книги, представляющие собой свод данных о редких, исчезающих и типичных растительных сообществах, нуждающихся в особой охране (Зеленая книга Сибири, 1996; Зеленая книга Украинской ССР, 1987г.).
В Красную книгу РСФСР, том «Растения», включено 516 видов растений и 17 видов (11 семейств) грибов. Растения представляют 440 видов (81 семейство) покрытосеменных, 11 видов (3 семейства) голосеменных, 10 видов (8 семейств) папоротниковидных, 4 вида (1 семейство) плауновидных, 22 вида (14 семейств) моховидных, 29 видов (13 семейств) лишайников. Данное издание до настоящего времени считается действующим, хотя и значительно устаревшим по многим позициям.
В настоящее время готовится новая версия Красной книги Российской Федерации и еще окончательно не определен список включенных видов растений и грибов. Поэтому в предварительном списке споровых растений и грибов были использованы данные Красного списка споровых растений и грибов (2004 г.) Государственного реестра охраняемых объектов животного и растительного мира России. Этот список включает 1263 таксона, из них 128 - папоротниковидные, 322 - лишайники, 91 - водоросли, 340 - грибы.
0 (Ех) - по-видимому, исчезнувшие виды (подвиды);
1 (Е) - виды (подвиды), находящиеся под угрозой исчезновения: таксоны, сохранение которых маловероятно, если факторы, вызвавшие сокращение их численности, будут продолжать действовать. К этой категории относятся таксоны, численность особей которых уменьшилась до критического уровня или число местонахождений которых сильно сократилось;
2 (V) - уязвимые виды (подвиды): таксоны, которым, по-видимому, в ближайшем будущем грозит перемещение в категорию находящихся под угрозой исчезновения, если факторы, вызвавшие сокращение их численности, будут продолжать действовать. К этой категории относятся таксоны, у которых численность особей всех или большей части популяций уменьшается вследствие чрезмерного использования, значительных нарушений или других изменений среды;
3 (Р) - редкие виды (подвиды), представленные небольшими популяциями, которые в настоящее время не находятся под угрозой исчезновения и не являются уязвимыми, но рискуют оказаться таковыми. Эти таксоны обычно распространены на ограниченной территории или имеют узкую экологическую амплитуду, либо рассеянно распространены на значительной территории;
4 (I) - виды (подвиды) с неопределенным статусом: таксоны, которые, очевидно, относятся к одной из предыдущих категорий, но достаточных сведений об их состоянии в настоящее время нет.
На карте показаны местонахождения наиболее редких видов сосудистых и споровых растений и грибов, включенных в Красную книгу РСФСР (1988 г.), имеющих статусы редкости 0(Ех) и 1(Е), а также местонахождения видов, опубликованных в Красном списке споровых растений и грибов России.

Выход в свет Красных книг СССР и РСФСР и разработка для них соответствующих законодательных актов послужили основой для появления региональных Красных книг. С середины 80-х годов прошлого столетия по настоящее время они изданы для 57 регионов, что составляет более половины общего числа субъектов Российской Федерации. Значение региональных Красных книг особенно возросло в 90-х годах прошлого столетия, когда в субъектах Российской Федерации повысилась роль самоуправления.
Помимо Красных книг, во многих регионах разработаны и большей частью утверждены региональные «Списки видов растений и животных, подлежащих охране». Обычно они представляют собой перечень русских и латинских названий видов, охраняемых в данном регионе, без указания на их распространение, численность и т.д. Многие официальные списки носят ведомственный характер, не опубликованы в широкой печати и до сих пор остаются практически недоступными для широкого круга заинтересованных лиц.
Число видов, включаемых в Красные книги отдельных регионов, варьирует от нескольких десятков до сотен. Из видов растений, включаемых в региональные Красные книги, наиболее многочисленны представители покрытосеменных растений. Некоторые книги ограничиваются только ими, но в большинство внесены также представители голосеменных, папоротниковидных, хвощевидных, плауновидных, моховидных, а из низших растений - водоросли и лишайники. Грибы в основном представлены видами из класса базидиальных грибов, но есть и представители слизевиков (Красная книга Ленинградской области).
Как правило, включенные в Красные книги России виды, распространенные на территории региона, включаются в региональные Красные книги. Но бывают и исключения, особенно в тех случаях, когда вид довольно обычен в регионе, и состояние его популяций не внушает опасений.
Одно из главных отличий растений от животных и грибов - способность создавать органические вещества из неорганических с помощью солнечного света (процесс фотосинтеза).Подцарство: Низшие растения
Тело низших растений (слоевище, или таллом) не разделено на истинные листья, стебель и корень, хотя может иметь их внешние подобия. Отделы (тип) :

Подцарство: Высшие растения
У высших растений тело разделяется на настоящие листья, стебель и корень. Отделы (тип):
- Моховидные, мхи, бриофиты
Наиболее примитивные из числа наземных растений. Встречаются в основном во влажных, затенённых местах. Мхи распространены во всех климатических поясах. У мхов нет настоящих проводящих тканей, вода и минеральные вещества поглощаются всей поверхностью тела. Высота не более 20 см. Большинство мхов - многолетние растения, встречаются группами (подушками, куртинками). Мохообразные - единственные наземные растения, у которых преобладает половое (гаплоидное) поколение - гаметофит. Бесполое поколение (спорофит) у мхов представлено спорогоном, который прикреплён к гаметофиту и питается за его счёт.- Папоротникообразные (споровые)
В основном наземные травянистые растения, встречаются также водные и древовидные формы. Предпочитают влажные и затенённые места.- Голосеменные
Голосеменные растения - древняя группа семенных растений, появившаяся в конце девона, около 370 млн. лет назад.Деревянистые растения.
Главное отличие от покрытосеменных (цветковых) - отсутствие цветков и плодов, а также сосудов и древесных волокон в стебле.
Семена лежат «голо», то есть не скрыты в завязи.
Голосеменные включают более 1 000 видов.- Покрытосеменные, цветковые
Наиболее высокоорганизованные наземные растения, травы, кустарники и деревья.
Основные отличительные признаки - наличие цветка и плода .
Семена скрыты (покрыты) в завязи, из которой образуется плод.
В стебле имеются сосуды и древесные волокна.
В настоящее время цветковые являются преобладающей формой наземной растительности
(описано свыше 250 тысяч видов).Класс:Наиболее крупные по числу видов семейства цветковых растений:
- Двудольные
В классе Двудольные описано 6 подклассов, 128 порядков, 418 семейств, приблизительно 10 000 родов и около 199 000 видов растений.
- зародыш семени имеет две семядоли,
- жилкование листьев - сетчатое,
- имеется центральный стержневой корень,
- количество лепестков и других частей цветка обычно кратно 4 или 5.
- Однодольные
Типичные отличительные признаки:В класс однодольных растений включаются 5 подклассов, 37 порядков, около 125 семейств, более 3000 родов и около 59 000 видов.
- зародыш семени имеет одну семядолю,
- жилкование листьев - параллельное или дуговое,
- корневая система - мочковатая,
- число частей цветка кратно 3.
- Астровые, или Сложноцветные (Asteraceae, Compositae) - 27773 видов в составе 1765 родов;
- Орхидные (Orchidaceae) - 27135 видов в составе 925 родов;
- Бобовые (Fabaceae, Leguminosae) - 23535 видов в составе 917 родов.
Грибы — древние гетеротрофные организмы, занимающие особое место в общей системе живой природы. Они могут быть как микроскопически малы, так и достигать нескольких метров. Поселяются на растениях, животных, человеке или на мёртвых органических остатках, на корнях деревьев и трав. Их роль в биоценозах велика и разнообразна. В цепи питания они являются редуцентами — организмами, питающимися мёртвыми органическими остатками, подвергающими эти остатки минерализации до простых органических соединений.
В природе грибы играют положительную роль: они пища и лекарства для животных; образуя грибокорень, помогают растениям всасывать воду; являясь компонентом лишайников, грибы создают среду обитания для водорослей.
Грибы — бесхлорофилльные низшие организмы, объединяющие около 100 000 видов, от мелких микроскопических организмов до таких великанов, как трутовики, гигантский дождевик и некоторые другие.
В системе органического мира грибы занимают особое положение, представляя отдельное царство, наряду с царствами животных и растений. Они лишены хлорофилла и поэтому требуют для питания готовое органическое вещество (принадлежат к гетеротрофным организмам). По наличию в обмене мочевины, в оболочке клеток — хитина, запасного продукта — гликогена, а не крахмала — они приближаются к животным. С другой стороны, способом питания (путём всасывания, а не заглатывания пищи), неограниченным ростом они напоминают растения.
Грибы имеют и признаки, свойственные только им: почти у всех грибов вегетативное тело представляет собой грибницу, или мицелий, состоящий из нитей — гиф.
Это тонкие, как нити, трубочки, заполненные цитоплазмой. Нити, составляющие гриб, могут туго или рыхло переплетаться, ветвиться, срастаться друг с другом, образуя плёнки наподобие войлока или видимые простым глазом жгуты.
У высших грибов гифы разделены на клетки.
В клетках грибов может быть от одного до нескольких ядер. Кроме ядер, в клетках имеются и другие структурные компоненты (митохондрии, лизосомы, эндоплазматическая сеть и пр.).

Строение
Тело подавляющего большинства грибов построено из тонких нитчатых образований — гиф. Совокупность их образует грибницу (или мицелий).

Разветвляясь, мицелий образует большую поверхность, что обеспечивает всасывание воды и питательных веществ. Условно грибы делятся на низшие и высшие. У низших грибов гифы не имеют поперечных перегородок и мицелий представляет собой одну сильно разветвлённую клетку. У высших грибов гифы разделены на клетки.
Клетки большинства грибов покрыты твёрдой оболочкой, её нет у зооспор и вегетативного тела некоторых простейших грибов. В цитоплазме гриба содержатся структурные белки и не связанные с органоидами клетки ферменты, аминокислоты, углеводы, липиды. Органоиды: митохондрии, лизосомы, вакуоли, содержащие запасные вещества — волютин, липиды, гликоген, жиры. Крахмала нет. В клетке гриба имеется одно или несколько ядер.
Размножение
У грибов различают вегетативное, бесполое и половое размножение.
Вегетативное
Размножение осуществляется частями мицелия, специальными образованиями — оидиями (образующимися в результате распадения гиф на отдельные короткие клетки, каждая из которых даёт начало новому организму), хламидоспорами (образуются примерно так же, но имеют более толстую тёмноокрашенную оболочку, хорошо переносят неблагоприятные условия), путём почкования мицелия или отдельных клеток.
Для бесполого вегетативного размножения специальные приспособления не нужны, но потомков появляется не много, а мало.
При бесполом вегетативном размножении клетки нити, ничем не отличаются от соседних, вырастают в целый организм. Иногда, животные или движение среды разрывают гифу на части.
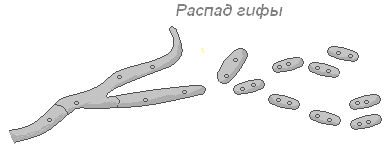
Бывает при наступлении неблагоприятных условий нить сама распадается на отдельные клетки, каждая из которых может вырасти в целый гриб.
Порой на нити образуются наросты, которые разрастаются, отпадают и дают начало новому организму.

Часто некоторые клетки наращивают толстую оболочку. Они могут выдерживать высыхание и сохраняют жизнеспособность до десяти и более лет, а в благоприятных условиях прорастают.

При вегетативном размножении ДНК потомков не отличается от ДНК родителя. При таком размножении не нужны специальные устройства, но количество потомков невелико.
Бесполое
При бесполом споровом размножении нить гриба образует специальные клетки, создающие споры. Эти клетки выглядят как веточки, неспособные расти и отделяющие от себя споры, или как крупные пузыри, внутри которых образуются споры. Такие образования называют спорангиями.
При бесполом размножении ДНК потомков не отличается от ДНК родителя. На образование каждой споры тратится меньше веществ, чем на одного потомка при вегетативном размножении. Бесполым путём одна особь производит миллионы спор, поэтому у гриба больше шансов оставить потомство.
Половое
При половом размножении появляются новые сочетания признаков. При этом размножении ДНК потомков образуется из ДНК обоих родителей. У грибов объединение ДНК происходит по-разному.
Разные способы обеспечить объединение ДНК при половом размножении грибов:



В какой-то момент сливаются ядра, а затем и нити ДНК родителей, обмениваются кусочками ДНК и разделяются. В ДНК потомка оказываются участки, полученные от обоих родителей. Поэтому потомок чем-то похож на одного родителя, а чем-то — на другого. Новое сочетание признаков может уменьшить, и увеличить жизнеспособность потомства.
Размножение состоит в слиянии мужских и женских половых гамет, в результате чего образуется зигота. У грибов различают изо-, гетеро- и оогамию. Половой продукт низших грибов (ооспора) прорастает в спорангий, в котором развиваются споры. У аскомицетов (сумчатых грибов) в результате полового процесса образуются сумки (аски) — одноклеточные структуры, содержащие обычно 8 аскоспор. Сумки образующиеся непосредственно из зиготы (у низших аскомицетов) или на развивающихся из зиготы аскогенных гифах. В сумке происходит слияние ядер зиготы, затем мейотическое деление диплоидного ядра и образование гаплоидных аскоспор. Сумка активно участвует в распространении аскоспор.
Для базидиальных грибов характерен половой процесс — соматогамия. Он состоит в слиянии двух клеток вегетативного мицелия. Половой продукт — базидия, на которой образуются 4 базидиоспоры. Базидиоспоры гаплоидны, они дают начало гаплоидному мицелию, который недолговечен. Путём слияния гаплоидного мицелия образуется дикариотический мицелий, на котором образуются базидии с базидиоспорами.
У несовершенных грибов, а в некоторых случаях и у других половой процесс заменяется гетерокариозом (разноядерностью) и парасексуальным процессом. Гетерокариоз состоит в переходе генетически неоднородных ядер из одного отрезка мицелия в другой путём образования анастомозов или слияния гиф. Слияние ядер при этом не происходит. Слияние ядер после, перехода их в другую клетку называется парасексуальным процессом.
Нити гриба прирастают поперечным делением (вдоль клетки нити не делятся). Цитоплазма соседних клеток гриба составляет единое целое — в перегородках между клетками есть отверстия.

Питание
Большинство грибов имеет вид длинных нитей, всасывающих питательные вещества всей поверхностью. Грибы всасывают нужные вещества из живых и мёртвых организмов, из почвенной влаги и воды природных водоёмов.
Грибы выделяют наружу вещества, разрывающие молекулы органических веществ на такие части, которые гриб может впитать.

Но в определённых условиях организму полезнее быть нитью (как гриб), а не комочком (циста) как бактерия. Проверим, так ли это.
Проследим за бактерией и растущей нитью гриба. Крепкий раствор сахара показан коричневым цветом, слабый — светло-коричневый, вода без сахара — белым.

Можно сделать вывод: нитевидный организм, разрастаясь, может оказаться в местах богатых пищей. Чем длиннее нить, тем больше запас веществ, который насытившиеся клетки могут расходовать на рост гриба. Все гифы ведут себя, как части одного целого, и участки гриба, оказавшись в богатых пищей местах, питают весь гриб.
Плесневые грибы
Плесневые грибы поселяются на увлажнённых остатках растений, реже животных. Одним из наиболее распространённых плесневых грибов является мукор, или головчатая плесень. Грибницу этого гриба в виде тончайших белых гифов можно обнаружить на залежавшемся хлебе. Гифы мукора не разделены перегородками. Каждая гифа представляет собой одну сильно разветвлённую клетку с несколькими ядрами. Одни ответвления клетки проникают в субстрат и поглощают питательные вещества, другие поднимаются вверх. На верхушке последних образуются чёрные округлые головки — спорангии, в которых образуются споры. Созревшие споры распространяются воздушными потоками или при помощи насекомых. Попав в благоприятные условия, спора прорастает в новую грибницу (мицелий).

Вторым представителем плесневых грибов является пеницилл, или сизая плесень. Грибница пеницилла состоит из гифов, разделённых поперечными перегородками на клетки. Некоторые гифы поднимаются вверх, и на конце их образуются разветвления, напоминающие кисточки. На конце этих разветвлений образуются споры, с помощью которых пеницилл размножается.

Дрожжевые грибы
Дрожжи — одноклеточные неподвижные организмы овальной или удлинённой формы, размером 8-10 мкм. Настоящего мицелия не образуют. В клетке имеется ядро, митохондрии, в вакуолях накапливается много веществ (органических и неорганических), в них происходят окислительно-восстановительные процессы. Дрожжи накапливают в клетках волютин. Вегетативное размножение почкованием или делением. Спорообразование наступает после многократного размножения почкованием или делением. Оно совершается легче при резком переходе от обильного питания к незначительному, при поступлении кислорода. В клетке число спор парное (чаще 4-8). У дрожжей известен и половой процесс.
Дрожжевые грибы, или дрожжи, встречаются на поверхности плодов, на содержащих углеводы растительных остатках. От других грибов дрожжи отличаются тем, что не имеют грибницы и представляют одиночные, в большинстве случаев овальные клетки. В сахаристой среде дрожжи вызывают спиртовое брожение, в результате которого выделяются этиловый спирт и углекислый газ:
С 6 Н 12 О 6 → 2С 2 Н 5 ОН + 2СО 2 + энергия.
Этот процесс ферментативный, протекает при участии комплекса ферментов. Освобождающаяся энергия используется дрожжевыми клетками на жизненные процессы.
Размножаются дрожжи почкованием (некоторые виды — путём деления). При почковании на клетке образуется выпуклость, напоминающая почку.

Ядро материнской клетки делится, и одно из дочерних ядер переходит в выпуклость. Выпуклость быстро растёт, превращается в самостоятельную клетку и отделяется от материнской. При очень быстром почковании клетки не успевают разъединяться и в результате получаются короткие непрочные цепочки.
Не менее ¾ всех грибов — сапрофиты. Сапрофитный способ питания связан преимущественно с продуктами растительного происхождения (кислая реакция среды и состав органических веществ растительного происхождения более благоприятны для их жизни).
Грибы-симбионты связаны преимущественно с высшими растениями, мохообразными, водорослями, реже — с животными. Примером могут быть лишайники, микориза. Микориза — это сожительство гриба с корнями высшего растения. Гриб помогает растению усваивать труднодоступные вещества гумуса, способствует поглощению элементов минерального питания, помогает своими ферментами в углеводном обмене, активизирует ферменты высшего растения, связывает свободный азот. От высшего растения гриб, очевидно, получает безазотные соединения, кислород и корневые выделения, способствующие прорастанию спор. Микориза очень распространена среди высших растений, она не обнаружена лишь у осоковых, крестоцветных и водных растений.
Экологические группы грибов
Почвенные грибы
Почвенные грибы участвуют в минерализации органического вещества, образовании гумуса и т.п. В этой группе выделяют грибы, попадающие в почву только в определённые периоды жизни, и грибы ризосферы растений, живущие в зоне их корневой системы.
Специализированные почвенные грибы:
- копрофиллы - грибы, обитающие на почвах, богатых перегноем (навозные кучи, места скопления помёта животных);
- кератинофиллы - грибы, обитающие на волосах, рогах, копытах;
- ксилофиты - грибы, разлагающие древесину, среди них различают разрушителей живой и мёртвой древесина.
Домовые грибы
Домовые грибы — разрушители деревянных частей построек.
Водные грибы
К ним относится и группа микоризных грибов-симбионтов.
Грибы, развивающиеся на промышленных материалах (на металле, бумаге и изделиях из них)
Шляпочные грибы
Шляпочные грибы поселяются на богатой перегноем лесной почве и из неё получают воду, минеральные соли и некоторые органические вещества. Часть органических веществ (углеводы) они получают от деревьев.
Грибница — главная часть каждого гриба. На ней развиваются плодовые тела. Шляпка и ножка состоят из плотно прилегающих друг к другу нитей грибницы. В ножке все нити одинаковы, а в шляпке они образуют два слоя — верхний, покрытый кожицей, окрашенной разными пигментами, и нижний.

У одних грибов нижний слой состоит из многочисленных трубочек. Такие грибы называют трубчатыми. У других нижний слой шляпки состоит из радиально расположенных пластинок. Такие грибы называют пластинчатыми. На пластинках и на стенках трубочек образуются споры, с помощью которых грибы размножаются.
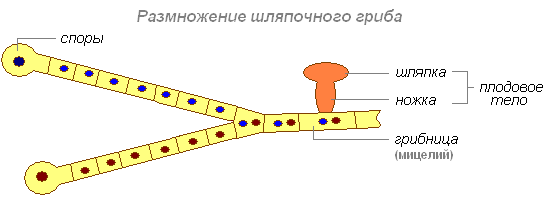
Гифы грибницы оплетают корни деревьев, проникают в них и распространяются между клетками. Между грибницей и корнями растений устанавливается полезное для обоих растений сожительство. Гриб снабжает растения водой и минеральными солями; заменяя на корнях корневые волоски, дерево уступает ему часть своих углеводов. Только при такой тесной связи грибницы с определёнными породами деревьев возможно образование плодовых тел у шляпочных грибов.
Образование спор
В трубочках или на пластинках шляпки образуются особые клетки — споры. Созревшие мелкие и лёгкие споры высыпаются, их подхватывает и разносит ветер. Разносят их насекомые и слизни, а также белки и зайцы, поедающие грибы. Споры не перевариваются в пищеварительных органах этих животных и выбрасываются наружу вместе с помётом.
Во влажной, богатой перегноем почве споры грибов прорастают, из них развиваются нити грибницы. Грибница, возникающая из одной споры, может образовывать новые плодовые тела лишь в редких случаях. У большинства видов грибов плодовые тела развиваются на грибницах, образованных слившимися клетками нитей, берущих начало от разных спор. Поэтому клетки такой грибницы двухъядерные. Грибница растёт медленно, лишь накопив запасы питательных веществ, она образует плодовые тела.
Большинство видов этих грибов — сапрофиты. Развиваются на перегнойной почве, отмерших растительных остатках, некоторые на навозе. Вегетативное тело состоит из гиф, образующих находящуюся под землёй грибницу. В процессе развития на грибнице вырастают зонтикоподобные плодовые тела. Пенёк и шляпка состоят из плотных пучков нитей грибницы.
У части грибов на нижней стороне шляпки от центра к периферии радиально расходятся пластинки, на которых развиваются базидии, а в них споры — это гименофор. Такие грибы называют пластинчатыми. У отдельных видов грибов имеется покрывало (плёночка из неплодных гиф), защищающее гименофор. При дозревании плодового тела покрывало разрывается и остаётся в виде бахромы по краям шляпки или кольца на ножке.
У некоторых грибов гименофор имеет трубчатую форму. Это трубчатые грибы. Их плодовые тела мясистые, быстро загнивают, легко повреждаются личинками насекомых, поедаются слизнями. Размножаются шляпочные грибы спорами и частями мицелия (грибницы).
Химический состав грибов
В свежих грибах вода составляет 84-94% общей массы.
Белки грибов усваиваются только на 54-85% — хуже, чем белки других растительных продуктов. Усвоению препятствует плохая растворимость белков. Жиры, углеводы усваиваются очень хорошо. Химический состав зависит от возраста гриба, его состояния, вида, условий произрастания и др.
Роль грибов в природе
Многие грибы срастаются с корнями деревьев и трав. Их сотрудничество взаимовыгодно. Растения дают грибам сахар и белки, а грибы разрушают находящиеся в почве мёртвые остатки растений и всасывают всей поверхностью гиф воду с растворёнными в ней минеральными веществами. Корни, сросшиеся с грибами, называют микоризой. Большинство деревьев и трав образуют микоризу.
Грибы играют в экосистемах роль разрушителей. Они уничтожают мёртвую древесину и листья, корни растений и трупы животных. Все мёртвые остатки они превращают в углекислый газ, воду и минеральные соли — в то, что могут усвоить растения. Питаясь, грибы набирают вес и становятся пищей животных и других грибов.
Растения делятся на две очень неравные по значению и величине группы – споровые и семенные растения.
К споровым растениям относятся отделы: мохообразные, псилотообразные, хвощеобразные, плаунообразные и папоротникообразные .
Отличительные признаки жизненного цикла споровых растений:
1. Размножаются спорами (семя никогда не образуется).
2. Процессы полового и бесполого размножения разделены (в пространстве и во времени). Причем, у большинства споровых растений, исключая мохообразные , половое (гаметофит) и бесполое (спорофит) поколения представляют собой отдельные физиологически самостоятельные особи.
3. В цикле развития у большинства споровых (за исключением мохообразных) спорофит преобладает (по размерам и сложности строения) над гаметофитом (который представлен заростком в виде слоевища (таллома) не расчлененного на стебель и листья, и тесно связан с водой).
4. Процесс оплодотворения у споровых растений, так же как и у водорослей, осуществляется в присутствии капельножидкой воды, так как сперматозоиды могут передвигаться только в воде.
5. Как равно-, так и разноспоровые растения (у некоторых равноспоровых, например, хвощей , мхов , существует физиологическая разноспоровость).
3. Общая характеристика семенных растений
Семенные растения – это растения, которые образуют семя.
К семенным растениям относится два отдела: голосеменные и покрытосеменные, которые возникли в процессе эволюции от споровых растений (разноспоровых папоротников).
По сравнению со споровыми они представляют собой более высокий уровень организации растений, так как:
1. Все семенные растения разноспоровые.
Они имеют два типа спор: микроспоры (дают начало мужскому гаметофиту) и мегаспоры (дают начало женскому гаметофиту). Гаметофит очень редуцирован и не выходит из споры, которая защищает его от высыхания, что является важным приспособлением к жизни на суше. Гаметафиты не способны к фотосинтезу и полностью зависят от спорофитов.
Женский гаметофит (мегагаметофит) заключен внутри мегаспоры (рис. 8.4.), которая образуется в мясистом мегаспорангии – нуцеллусе, расположенном на мегаспорофилле (видоизмененном спороносном листе). В отличие от бессемянных разноспоровых растений мегаспорангий покрыт одним или двумя дополнительными слоями ткани – интегументами. Они полностью заключают в себя мегаспорангий (нуцеллус), оставляя лишь отверстие на его верхушке – микропиле (пыльцевход).
Рис. 8.4. Строение семязачатка (схема).
Прорастание мегаспоры и образование женского гаметофита (мегагаметофита), оплодотворение и развитие нового спорофита (зародыша) всегда происходит внутри мегаспорангия.
2. Образование семян.
У семенных растений (в отличие от споровых) мегаспоры не отделяются от спорофита, а находятся внутри мегаспорангия, который вместе с интегументами представляет семязачаток. Внутри мегаспоры развивается женский гаметофит (зародышевый мешок) и образуется одна или несколько женских гамет – яйцеклеток. После оплодотворения яйцеклетки семязачаток превращается в семя. Таким образом, семя – это оплодотворенный семязачаток.
Семя состоит из: зародыша будущего растения (корешок и почечка (стебелек и листочки )), который возникает из оплодотворенной яйцеклетки; определенного количества питательных веществ; кожуры, которая возникает из интегументов.
В борьбе за существование на суше появление семени имело большое эволюционное значение, поскольку внутри него (в отличие от споры) уже находится зародыш нового спорофита, который надежно защищен от воздействия неблагоприятных условий. Также семя имеет существенное преимущество при размножении перед спорой, так как содержит значительный запас питательных веществ. У семян могут развиваться различные приспособления, облегчающие их распространение.
Родительский спорофит дает семени все, что нужно для жизни, и только после того, как семя полностью созреет, оно отделяется от родительского спорофита.
3. Независимое от воды оплодотворение.
Мужские гаметы (у большинства семенных – спермии, у некоторых голосеменных (саговниковые, гинкговые) – сперматозоиды) образуются мужскими гаметофитами (микрогаметофитами), находящимися внутри пыльцы. Пыльца из микроспорангиев (пыльцевых мешков) переносится к семязачаткам. Такой процесс называется опылением.
Пыльца прорастает пыльцевой трубкой , которая растет в сторону яйцеклетки (+ хемотаксис). По этой трубке неподвижные мужские гаметы (спермии) достигают яйцеклетки и происходит оплодотворение.
Вода не нужна спермиям ни на одной из перечисленных стадий!
Гаметофитное поколение у семенных растений крайне редуцировано (особенно у покрытосеменных), теряет свою самостоятельность (сравните со споровыми растениями) и развивается на спорофите, который сильно усложняется (как и у всех сосудистых растений доминирует в жизненном цикле).
Внутреннее оплодотворение, независимое от воды, развитие зародыша внутри семени, представляют собой главное биологическое преимущество семенных растений, которое позволило им освоить разнообразные местообитания и стать господствующей группой растений на Земле.